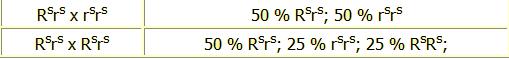| 
Genetikai alapfogalmak
A sejtek kromoszómaszáma egy adott fajra jellemző, állandó érték. A kromoszómákban (DNS-molekulákban) találhatók a gének, ezek az öröklődés egységei. A gének kódolt utasításokat tartalmaznak, molekuláris szinten lehetővé teszik, hogy az élő szervezet előállíthassa a fejlődéséhez, növekedéséhez, működéséhez és szaporodásához szükséges anyagokat (polipeptideket, fehérjéket, enzimeket stb.).
A génváltozatok, az allélek az élőlények változatosságát okozzák, meghatározzák az egyedek tulajdonságait. Allélnek nevezzük a homológ kromoszómákon, azonos helyen (lokuszon) található gének egyik lehetséges változatát. Számos lokusz több különböző allélt hordozhat, ezek egy változó tulajdonságot határoznak meg.
Genotípus: egy adott szervezet génszerelvénye.
Fenotípus: egy szervezet minden olyan jellege, amely genetikai teszt nélkül meghatározható- ilyenek a látható jellegek, amelyek a genetikai és környezeti hatások kölcsönhatása révén jönnek létre.
Homozigóta jelenség: Két azonos kromoszóma azonos alléleket foglal magába, vagyis egy génnek két azonos tulajdonságot meghatározó allélje van.
Heterozigóta jelenség: Két azonos kromoszóma eltérő alléleket foglal magába, egy génnek két különböző tulajdonságot meghatározó allélje van.
Domináns allél: felerősíti az általa meghatározott tulajdonság hatását (nagybetűvel jelöljük).
Recesszív allél: azonos kromoszómában elhelyezkedő hasonmás gén, amely a domináns allél jelenlétében nem fejti ki hatását. A recesszív gén csak akkor juthat érvényre, ha mindkét homológ kromoszómában egyszerre van jelen (kisbetűvel jelöljük).
P (Parents): Szülői nemzedék.
F1 nemzedék: A szülőktől származó utódok.
F2 nemzedék: Azok az utódok, amelyek az F1 nemzedék egyedeinek kereszteződése által jöttek létre.
Melanin: a testben képződő pigment. Anyagcsereútja során a tirozin aminosavból a DOPA és ebből a dopakinon keletkezik (mindkét lépést a tirozináz enzim katalizálja). A továbbiakban polimerizáció révén alakul ki a fekete vagy barna eumelanin, amely fehérjékkel alkotott komplex a melanoszómákban. A vörös vagy sárga feomelaninok úgy jönnek létre, hogy a dopakinon ciszteinnel kapcsolódik, mielőtt az említett fehérjekomplexbe lépne.
A tengerimalac testi (szomatikus) sejtjei, a magasabb rendű élőlényekéhez hasonlóan diploidok, vagyis kétszeres kromoszómakészletű sejtek. A tengerimalac diploid kromoszómaszáma 2n=64, melyből 62 autoszóma (ezek testi kromoszómák, vagy más néven szomatikus kromoszómák) 31 homológ párt alkot, a fennmaradó 2 ivari kromoszóma (nemi kromoszóma) határozza meg az állat nemét, az xx a nőnemet (két homológ x kromoszóma), az xy pedig a hímnemet (eltérő felépítésű x és y kromoszómák). Az ivarsejtek kromoszómaszáma a testi sejtekének a fele. A petesejt és a hím ivarsejt haploid, vagyis egyszeres kromoszómakészletű sejt, amelyben nincsenek homológ kromoszómák. A tengerimalac ivarsejtjeiben 32 kromoszóma található.
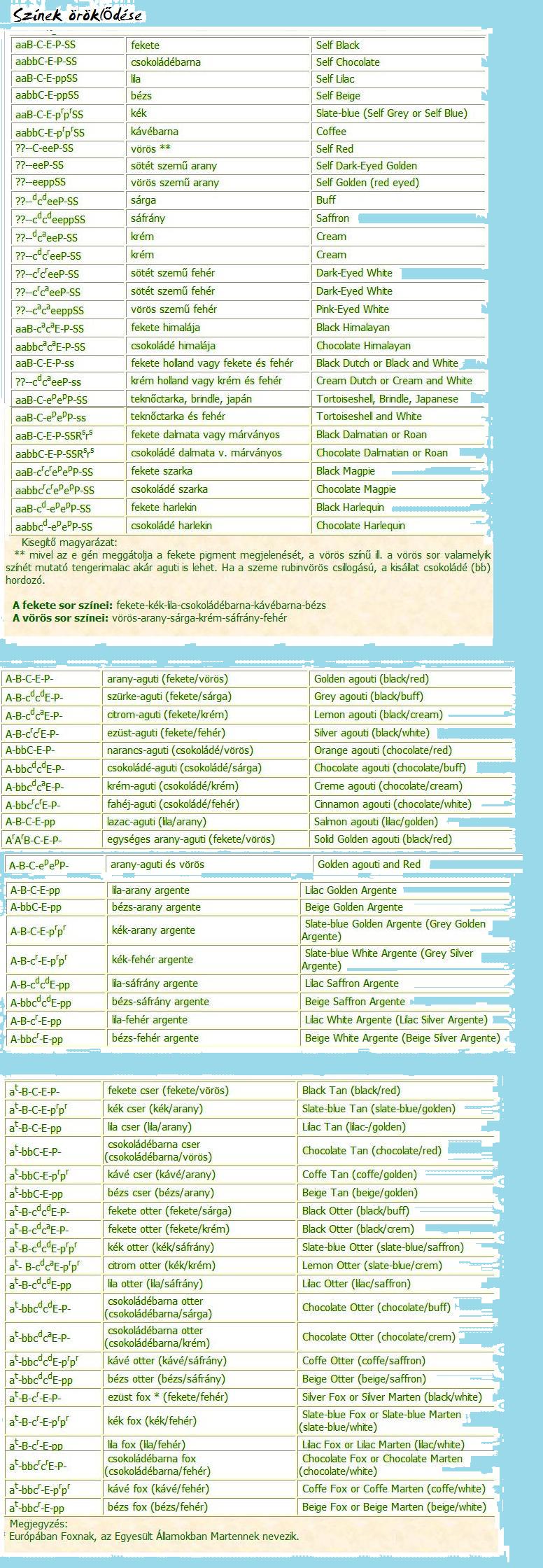
SZÍNVÁLTOZATOK ÖRÖKLŐDÉSE
A lokusz - nevezhetjük aguti faktornak is (Agouti locus).
A gén az eumelaninnak és feomelaninnak egy adott szőrszálon belüli eloszlását befolyásolja.
Eumelanin - fekete vagy barna (black or brown) pigment.
Feomelanin (Phaeomelanin) - sárga és vörös (yellow and red) pigment.
A - aguti allél (light-bellied agouti allele): A Cavia porcellus vad ősének színe az arany-aguti szín. A különböző aguti színek az arany-aguti szín módosulásai, minden egyes szőrszál terminális vagy szubterminális sávja a feomelanin által meghatározott, vagy ennek hígított változata, a fennmaradó rész pedig fekete vagy barna (eumelanin), vagy ezek egyikének hígított változata. A hasán a szőrszálak egyszínűek, a test egyéb részein található szőrszálak terminális vagy szubterminális sávjának megfelelő színűek. Az aguti színről részletesebb információ a tengerimalac-fajták oldalon található.
Ar - szolid aguti allél, egységes aguti (solid agouti, no belly band): Minden egyes szőrszál aguti színű, a szolid-agutinak (egységes agutinak) a hasán is aguti színűek a szőrszálak.
a - nem aguti allél (self, non-agouti allele): ez az allél minden egyes szőrszálról eltávolítja a feomelanin sávot, így a tengerimalac fekete vagy barna színű (vagy egy másik gén által meghatározott színű) lesz.
Domináns-recesszív öröklődésmenet - A dominancia sorrendje: A, Ar, a.
AA allélpár - a tengerimalac aguti színű.
Aa allélpár - a tengerimalac aguti színű, mert az a allél nem fejti ki hatását a domináns allél jelenlétében, de a későbbi nemzedékben kifejeződhet.
aa allélpár - nem aguti.
B lokusz - barna faktor (brown locus).
B - vad típusú allél: fekete szín (wild type allele: black colour)
b - barna allél (brown allele): A b allél a tengerimalac szőrszálainak, bőrének és szemének színére van hatással, így azok barna (csokoládébarna) színűre hígulnak. Az allél csak a fekete színre van hatással, a vörösre nincs.
Domináns-recesszív öröklődésmenet:
BB allélpár - a tengerimalac szőre fekete színű.
Bb allélpár - a tengerimalac szőre fekete színű, mert a b allél nem fejti ki hatását a domináns allél jelenlétében, de utódaiban megjelenhet a csokoládé szín.
bb allélpár - nem fekete, a tengerimalac szőrszála barnára, vagyis csokoládészínűre hígul, a szem rubinvörös csillogású.
C lokusz - a szín intenzitását meghatározó faktor (colour intensity locus).
Számos mutáns allél ismert, ezek mindkét színt (fekete, vörös) hígítják, a vörös pigmentre azonban nagyobb hatással vannak, mint a feketére. Itt számos helyzetben részleges dominancia jelentkezik. A krém szín intermedier a sárga és fehér között (a két allél gén egymással egyenlő erővel lép fel).
C allél - Vad típusú allél, fekete és/vagy vörös (ilyen az arany-aguti, a fekete, és a vörös szín) a szem sötét színű (wild type allele).
ck allél - a fekete és/vagy vörös színt hígítja (dark dilution of black and/or red)
cd allél - a fekete és/vagy vörös pigmentet kissé erőteljesebben hígítja (light dilution of black and/or red)
cr allél - a szemet rubinvörös színűre, a fekete pigmentet világos-szépia színűre, a vörös pigmentet pedig fehérre hígítja (csincsilla-faktornak is nevezik).
ca allél - homozigóta recesszív formában a himalája megjelenítésért felelős, a vörös pigmentet fehérre hígítja, a fekete pigmentet a „végekre” (orr, fül, végtagok) korlátozza. Az írisz és retina pigmenthiánya miatt a szem vörös színű lesz. A sötét szín kifejlődése a hőmérséklettől függ, a pigmentet előállító enzimnek az alacsonyabb hőmérsékleti értékek kedveznek, minél alacsonyabb a hőmérséklet, annál sötétebb a szín, és annál kiterjedtebbek a sötét területek. Egyesek cah vagy ch allélként is jelölik.
Néhány példa - allélikus kölcsönhatások:
Megj. - A fekete színnél:
- BB vagy Bb allélpár esetén a fekete szín hígulásai
- bb allélpár esetén a barna, vagyis a csokoládészín hígulásai
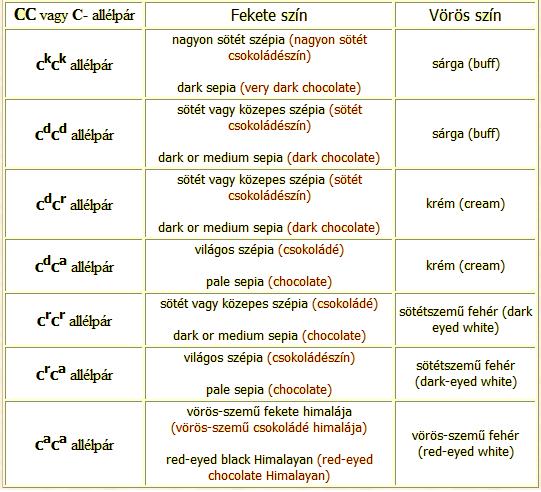
Megjegyzés: a C- allélpárnál a - jel a ck, cd, cr vagy ca allélok bármelyike lehet. A vad típusú allél jelenlétében a mutáns allél nem fejti ki hatását.
Fontos: Tudnunk kell, hogy a fehér tengerimalac nem albinó (pl. a fehér egerek és patkányok albinók, mert ezeknél a melanin pigment képzéséért felelős enzim hiányzik), így nevezik azt a tengerimalacot, amelynek szőre vörös lenne az E lokusz ee allélpárja miatt, de a caca allélpár a vörös pigment megjelenését gátolja (Dunkin-Hartley „albino” tengerimalacként ismert, mert Dunkin és Hartley közös laboratóriumában jelentek meg először).
E lokusz - Az E faktor az eumelanin és a feomelanin közötti viszonyt, a színek teljes testfelületen való kiterjedését befolyásolja (extension gene).
E allél - a teljes testfelületre a fekete- vagy barna szín (csokoládé) terjed ki, vagy ezek hígulásai (lila, bézs) jelennek meg, a tengerimalacon nincs vörös szín, sem ennek hígulásai (full extension).
ep allél - ez az allél a fekete szín kiterjedését csak foltok formájában engedélyezi, így a test teljes felületén foltokban jelenik meg a fekete, vagy annak hígított változata, és a vörös, vagy annak hígított változata (partial extension). Ez az allél felelős a teknőctarka, brindle és japán megjelenítésért. Sok kis gén befolyásolja az allélek hatásait, a foltok nagyságát és helyét, így rengeteg változat adódhat.
e allél - meggátolja a fekete pigment megjelenítését, csak a vörös, vagy ennek hígított változata (arany, sáfrány, sárga, krém vagy fehér) jelenik meg (non extension).
Az ep allél inkomplett domináns az e allélel szemben.Allélikus kölcsönhatások:

P lokusz - A P faktor a szem színét befolyásolja, az eumelaninre, vagyis a fekete/barna pigmentre van hatással (pale factor).
A P és B jellegeket kialakító gének egy kromoszómán, egymáshoz közel helyezkednek el, így a két tulajdonság nem függetlenül, hanem kapcsoltan öröklődik.
Domináns-recesszív öröklődésmenet:
P allél - nincs hígulás, a tengerimalac szeme sötét, szőre fekete színű (dark eyes).
pr allél - kevésbé ismert gén, a szem rubinvörös lesz, a fekete szín pedig palakékre hígul. A szőrszínt egyesek kéknek, palakéknek, mások szürkének nevezik (blue, slate-blue, gray). Svédországban az allél jelölése pg.
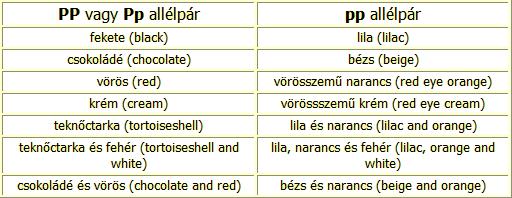
p allél - a szem vörösre hígul, a fekete szín lilára, a csokoládé bézsre (pink eye dilution gene).
A pp allélpár jelenléte nemcsak a fekete pigmentre, a vörös pigmentre is hatással van, a következő táblázat megmutatja, hogyan:
S lokusz - Az S faktor a fehér szín megjelenítéséért felelős (white spotting factor).
SS allélpár - nem jelenik meg a fehér szín.
Ss allélpár - inkomplett dominancia, a tengerimalac testén nagyon kicsi arányban megjelenhet a fehér szín, pl. a lábvégeken, az orron.
ss allélpár - a tengerimalac testén sok a fehér szín.
Sok kis gén befolyásolja az allélek hatásait, ezért a fehér megjelenésének aránya mindig változó.
Érdekesség: Az s allél másodlagos hatása egyéb színekre - amellett, hogy megjeleníti a fehér színt, a meglévő színeket is arra „kényszeríti”, hogy azok foltokban jelenjenek meg. Egy jó példa erre a teknőctarka és fehér tengerimalac: nem lehet brindle és fehér (mert a színek nem jelennek meg egyenletes eloszlásban), vagy japán és fehér (mert a színek nem jelennek meg csíkokban).
Dalmata és márványos faktor Az Rs gén Rsrs allélpár esetén a dalmata és márványos szín megjelenítéséért felelős (dalmatian or spotted roan faktor). A gén egy mutáció során jött létre, és a színelosztást befolyásolja. Az rs allél itt egy színes, egyszínű tengerimalac-szülőtől származó allélt jelent. A dalmata tengerimalacot egyes szakirodalomban Rnrn-ként is jelölik.
Fontos!
Dalmata vagy márványos tengerimalacot nem szabad maguk közt párosítani, mert a gén homozigóta domináns formában életképtelen tengerimalac megjelenését okozza. Letális génnek nevezik, mert sejtszinten a gén olyan változást idéz elő, amely a tengerimalac fejlődését korlátozza, így rendellenességgel születik, általában fehér színű (ezért Lethal Whitenak is nevezik), szeme kicsi és/vagy deformált, nincs foga, vagy ez is deformált, esetleg süket vagy steril, és legtöbbször néhány hetes korában elpusztul.
Rsrs allélpár - A dalmata tengerimalacnál a meglévő színt a fejre, a lábakra, a testen pedig pöttyökre korlátozza, a test többi része fehér (az orrán végighúzódik egy fehér sáv). A márványos tengerimalac abban különbözik a dalmatától, hogy a törzsén a pöttyök helyett egyenletesen keverednek a fehér és színes szőrszálak.
RsRs allélpár - letális gén. Az ilyen tengerimalac legtöbbször torzszülött vagy halva születik.
rsrs allélpár - az egyszínű (színes, nem fehér) tengerimalacot jelöljük így. A dalmata vagy márványozott tengerimalacot egyszínűvel érdemes párosítani, leginkább fekete vagy csokoládészínűvel.
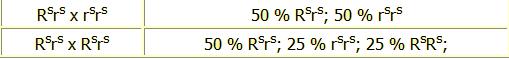
Kevésbé ismert a Cser (Tan) tengerimalac színét meghatározó mutáns gén.
Az arany-agutihoz hasonló, azzal a különbséggel, hogy minden egyes szőrszála csak egyszínű. Testén a szőr fekete vagy a fekete sor egyik színe, a szőrszálakon itt nincs sávonkénti színelosztás, mint az agutinál, a hasán lévő szőrzet pedig vörös színű vagy ennek egyik színe. A hígított változatokat Foxnak (az USAban Martennek) és Otternek nevezik. A cser tengerimalacoknál a színkombinációk ugyanazok, mint az aguti és argente változatoknál.
SZŐRVÁLTOZATOK ÖRÖKLŐDÉSE
A szőr hosszát meghatározó faktor:
L allél - rövidszőrű (Shorthair)
l allél - hosszúszőrű (Longhair)
LL allélpár - a tengerimalac szőre a test minden részén egyenletesen rövid.
Ll allélpár - az L allél inkomplett domináns az l allélel szemben, a tengerimalac szőre félhosszú, vagy bizonyos testrészein hosszabb szőrtincsek találhatók (oldalszőr, uszály).
ll allélpár - a tengerimalac szőre hosszú.
A bóbitát meghatározó faktor (Star gene):
St allél - bóbitás (Crested). A fejtetőn megjelenő rozettát nevezzük bóbitának.
Az angol bóbitás tengerimalac (English Crested) szőre egyszínű, a fejtetőn lévő rozetta a testével azonos színű.
Az amerikai bóbitás (American Crested) szőre kétszínű, teste színes, a bóbita pedig kötelezően fehér.
st allél - nem bóbitás (Non-Crested)
Domináns-recesszív öröklődésmenet:
StSt allélpár - bóbitás tengerimalac.
Stst allélpár - a tengerimalac bóbitás, a domináns allél jelenlétében az st allél nem fejti ki hatását, de a későbbi nemzedékben (homozigóta recesszív formában) kifejeződhet.
stst allélpár - nem bóbitás.
A rozettákat (forgókat) meghatározó faktor:
Rh m allél - rozettás (Rosette). Az Rh gén a forgók megjelenítéséért felelős, és 4 lokusz által szabályozott. Az m a rozetták helyét, alakját és/vagy számát módosító gén, feltételezhetően számos kis módosító gén is befolyásolja az allélek hatásait.
rhrh MM - simaszőrű (nem rozettás) tengerimalac.
Rh - domináns gén
M - inkomplett domináns gén, a rozetták helyét, alakját, számát csökkentő és/vagy gátló gén *.
rh - recesszív gén
* Megjegyzés: Kiváló rozettákkal rendelkező abesszin az RhRh mm tengerimalac. Az RhRh Mm, RhRh MM, Rhrh Mm vagy Rhrh MM tengerimalac szőrében a rozetták formája, száma, helye módosul vagy szőrzetében ridge (taréj) alakul ki (szimetrikus, láthatóan határolt, az állat egyéb testrészein található szőrzethez viszonyítva hosszabb, ellentétes irányba növő szőrzet végig a háton, általában simaszőrű-perui párosításnál).
Az Rh m allélpárok homozigóta formában az Abesszin szőrváltozatot jelenítik meg. Az abesszin tengerimalac szőrén általában 8 forgó található. A rozetták helye: 4 forgó a test törzsén, 2 a csípőn, 2 pedig a faron van. Ideális esetben további 1-1 forgó található az orra közelében, és szintén 1-1 a vállakon. Az abesszin tengerimalacnál tehát legalább 8 rozetta a követelmény (kiváló, ha 10 rozettával rendelkezik). Tudnunk kell, hogy a test törzsén található forgók megszabott helye és elrendezése mindig fontosabb, mint a 8-nál több rozetta.
A törzsön lévő 4 rozetta a test közepén, ennek mindkét oldalán szimmetrikusan, egyenes vonalban helyezkedik el. A csípőn és a faron lévő 2-2 rozetta is mindkét oldalon arányos, és a törzsön található forgókkal párhuzamos. A vállakon a forgók az elülső lábak fölött találhatók (mindkét oldalon 1-1), ezek kisebbek - az orra közelében lévő forgókkal megegyező nagyságúak. Az utóbbi 4 rozetta nem követelmény, de igen előnyős, mert megnyerő külsőt kölcsönöz az abesszin fajtának. A kb. 3, 5 cm szőrhossz a megfelelő, ennél rövidebb vagy hosszabb szőr esetén a rozetták mélysége és alakja nem lesz megfelelő.
Ha abesszin tengerimalacot simaszőrűvel párosítunk, az utódok szőrében általában 8-nál kevesebb rozetta jelenik meg. Az M inkomplett domináns gén hatására a rozetták formája, helye és/vagy száma módosul.
A rex-szőrminőséget meghatározó faktor:
A Rex (európai rex) szőrváltozat Angliában jelent meg.
A rex szőrzetet kialakító gén recesszív, csak akkor jut kifejeződésre, ha mindkét homológ kromoszómában egyszerre van jelen.
RxRx allélpár - nem-rexoid gén, a tengerimalac szőre sima (Non-Rexoid, no curly hair)
Rxrx allélpár - a tengerimalac szőre sima, a domináns allél jelenlétében a recesszív allél nem fejti ki hatását
rxrx allélpár - rexoid szőrminőségű tengerimalac (Rexoid gene, curly hair).
Hasonló szőrminőségű az amerikai rex-fajta, ez Teddy néven ismert, az Egyesült Államokban jelent meg.
FzFz allélpár - nem-rexoid gén, a tengerimalac szőre sima
Fzfz allélpár - a tengerimalac szőre sima, a domináns allél jelenlétében az fz allél nem fejti ki hatását.
fzfz allélpár - Teddy (amerikai rex)
Fontos! Habár azonos fenotípusúak, genotípusuk eltérő, ezért e két fajtát nem ajánlatos egymás közt szaporítani. Ha Rex tengerimalacot Teddyvel párosítunk, az F1 nemzedékben az utódok simaszőrűek lesznek.
rxrx FzFz - Rex (európai rex)
RxRx fzfz - Teddy (amerikai rex)

Megemlítendő:
Szintén rex-szőrminőségű a Curly fajta, amely Svédországban (Stockholmban) jelent meg először. A Rex, Teddy és Curly tengerimalacok genetikailag különbözőek, csak fenotípusuk hasonló. Mindhárom gén egymástól függetlenül, spontán mutáció során jött létre.
A curly szőrzetet meghatározó gén domináns, a Rex és Teddy recesszív. Peruival párosítva jelent meg a Lunkarya fajta.
Szatén szőrváltozat (Satin):
Erős csillogású, különlegesen fényes szőrzet, bármely színben és szőrtípusban megjelenhet. A szatén tengerimalac egyes szőrszálai üregesek, szőrének csillogása ennek köszönhető.
A szatén szőrzetet meghatározó gén recesszív, csak akkor jut kifejeződésre, ha mindkét homológ kromoszómában egyszerre van jelen.
Domináns-recesszív öröklődésmenet:
SnSn allélpár - nem szatén (normál szőrzetű) tengerimalac.
Snsn allélpár - nem szatén tengerimalac (más szóval szatén-örökítő), a domináns allél jelenlétében az sn allél nem fejti ki hatását, de a későbbi nemzedékben kifejeződhet.
snsn allélpár - szatén tengerimalac.
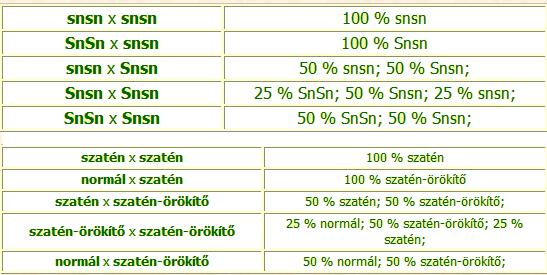
A szőrnélküliséget (kopaszságot) meghatározó faktor:
A kopaszságot meghatározó jelleg recesszív, csak akkor jut kifejeződésre, ha a szervezet homozigóta az adott génre. A gén spontán mutáció eredménye. Kutatóintézetek laboratóriumaiban jelent meg először, s később magántenyészetekbe is eljutott. Színes szőrős tengerimalacokkal párosítva jelentek meg a különböző színű kopasz tengerimalac-változatok.
A kopaszságot meghatározó jelleg recesszív, csak akkor jut kifejeződésre, ha a szervezet homozigóta az adott génre. A gén spontán mutáció eredménye. Kutatóintézetek laboratóriumaiban jelent meg először, s később magántenyészetekbe is eljutott. Színes szőrős tengerimalacokkal párosítva jelentek meg a különböző színű kopasz tengerimalac-változatok.
Különleges bánásmódot igényelnek, nagyon tiszta almot, az átlagosnál magasabb környezeti hőmérsékletet, a szokásosnál több táplálékot - mindezt éppen azért, mert nincs bundájuk, érzékenyebbek a hidegre, több kalóriát fogyasztanak el a test hőmérsékletének fenntartására.
Két fajtát ismerünk:
Hairless, vagy más néven Skinny tengerimalac (szőrnélküli)
Nem teljesen kopasz, az orr körül és a lábvégeken néhány szőrszál található - a szőrszálak mennyisége és kiterjedése egyénenként változó, egyeseknél több, másoknál kevesebb.
Az első példányok 1968-ban, Angliában, Millhill városában jelentek meg, a helyi orvosi kutatóintézetben (Medical Research Council). Ezek vörösszemű fehér tengerimalacok (Hartley „albino” tengerimalacok) utódai voltak.
Szőrnélküli tengerimalacok 1978-ban, Kanadában, Montreal városában is megjelentek, az „Armand Frappier” intézetben (Institut Armand Frappier). E mutáció beltenyészetben keletkezett, három nőstény és egy hím utódainál. Ezek vörösszemű fehér tengerimalacok (szintén Hartley „albino” tengerimalacok) voltak.
Baldwin (kopasz)
A Baldwin-fajta viszonylag kevésbé elterjedt. 1994-ben, az Egyesült Államokban (San Diegoban) jelent meg. A mutáció egy magántenyészetben, két aranyszínű amerikai bóbitás (Golden American Crested) utódainál keletkezett. Szőrösen jöttek a világra, majd lassan, fokozatosan vesztették el szőrzetüket, kb. 3 hét leforgása alatt váltak teljesen kopasszá. A szülők újabb szaporítása nyomán az első alomhoz hasonló (szőrüket fokozatosan elvesztő, majd kopasszá váló) utódok születtek.
A Baldwin-fajta teljesen kopasz, bőre (a szőrnélkülihez viszonyítva) vékonyabb, és általában nincs bajusza, szempillája.
A Skinny és Baldwin bőre enyhén ráncosodó.
Domináns-recesszív örökölődésmenet:

Fontos!
* Csak Skinny x Skinny, Baldwin x Baldwin esetében érvényes. Ha a két, genetikailag különböző fajtát egymás között párosítjuk, az utódok (F1 nemzedék) általában szőrős tengerimalacok lesznek.
normál - nem kopasz tengerimalac (domináns allél).
kopasz-örökítő - normál szőrzetű tengerimalac, heterozigóta. A domináns allél jelenlétében a kopaszság (szőrnélküliség) jelleg nem jut kifejezésre, de a későbbi nemzedékben megjelenhet.
kopasz - vagyis szőrnélküli tengerimalac (recesszív allél).
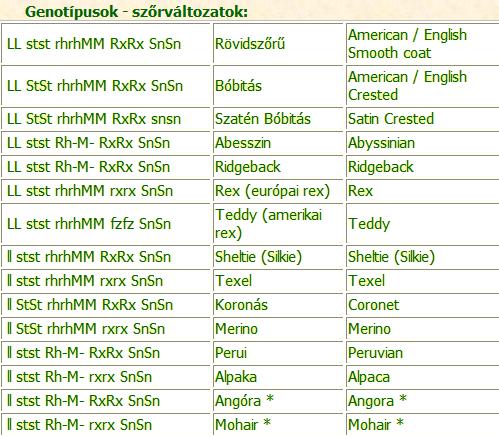
* a rozetták számának módosulásával (with a modifer for the number of rosette).
Az angóra a peruihoz hasonló, de ennél több rozettával rendelkező tengerimalac.
A mohair az alpakához hasonló, de ennél több rozettával rendelkező tengerimalac.
********************************
A tengerimalac genotípusának ismerete nagyon fontos a tenyésztésnél.Csupán a fenotípus ismeretével, nem tudhatjuk pontosan milyen utódokra számíthatunk. Genotípusára ilyenkor a leszármazottak tulajdonságai alapján következtethetünk.
forrás: www.gemon.ro
| |